
Planetary Scale Information Transmission in the Biosphere and Technosphere: Limits and Evolution


1
Department of Aerospace, Physics and Space Sciences, Florida Institute of Technology, Melbourne, FL 32901, USA
2
Department of Physics and Institute for Fusion Studies, The University of Texas at Austin, Austin, TX 78712, USA
3
Department of Physics and Astronomy, University of Rochester, Rochester, NY 14620, USA
4
Dipartimento di Fisica, Università di Roma “Tor Vergata”, 00133 Roma, Italy
*
Author to whom correspondence should be addressed.
Life 2023, 13(9), 1850; https://doi.org/10.3390/life13091850
Submission received: 19 July 2023
/
Revised: 24 August 2023
/
Accepted: 29 August 2023
/
Published: 31 August 2023
(This article belongs to the Section Astrobiology)
{kind=link}
Abstract
:Information transmission via communication between agents is ubiquitous on Earth, and is a vital facet of living systems. In this paper, we aim to quantify this rate of information transmission associated with Earth’s biosphere and technosphere (i.e., a measure of global information flow) by means of a heuristic order-of-magnitude model. By adopting ostensibly conservative values for the salient parameters, we estimate that the global information transmission rate for the biosphere might be ∼ bits/s, and that it may perhaps exceed the corresponding rate for the current technosphere by ∼9 orders of magnitude. However, under the equivocal assumption of sustained exponential growth, we find that information transmission in the technosphere can potentially surpass that of the biosphere ∼90 years in the future, reflecting its increasing dominance.
1. Introduction
It is widely accepted that information processing constitutes a fundamental aspect of life, e.g., [1,2,3,4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20]. In this “informational” realm, the significance and ubiquity of information transmission between organisms/agents via appropriate communication channels is thoroughly documented, ranging from microscopic to macroscopic scales and spanning multiple modalities, e.g., [21,22,23,24,25,26,27,28,29,30,31].
A number of authors have hypothesized that exploring information transmission via communication might be valuable for understanding various major evolutionary events in Earth’s history. The proposals have ranged from abiogenesis and early evolution of (proto) life [22,28,31,32,33,34] to breakthroughs in cooperation, collective behavior, multicellularity and cognition [18,30,35,36,37,38,39,40]. Even setting aside these concepts, comparisons of key properties of Earth’s biosphere and “technosphere” (see, e.g., [41,42]) can yield potentially useful insights into how much the latter is catching up with, or impinging, on the former, which is increasingly timely in the Anthropocene [43,44]. Comparisons of aspects of the biosphere and the technosphere have recently been published in the context of mass [45] and internal information processing [46].
Hence, it is natural to wonder about the aforementioned rate of information transmission (measured in bits/s) ascribable to modern Earth’s biosphere and its technosphere, at a given instant in time, which follows from generalizing this notion to encompass the planet in toto—the overarching theme of this paper. Analyzing the global rate of information transfer may shed light on the communication load embedded in planetary-scale biological and technological systems. This theme is important both from the standpoint of understanding the genesis and evolution of these systems on Earth, as well as assessing the astrobiological possibility of their emergence elsewhere [47,48,49,50].
Due to the manifold uncertainties at play (as illustrated hereafter), we report a simple order-of-magnitude estimate (Fermi problem) in the spirit of the famous Drake equation from astrobiology [51,52,53]. We will first examine the global rate of information transmission in the biosphere (Section 2), and then delve into the equivalent calculation for the human-mediated technosphere (Section 3). Finally, we articulate the ramifications of our preliminary findings in Section 4.
2. Information Transmission in the Biosphere
We shall focus on prokaryotes in this analysis (denoted by the subscript ‘p’), whose communication is modulated by signaling molecules [54,55]. The total biomass of eukaryotes is probably times greater than that of prokaryotes ([56], Figure 1), but the mean volume, and consequently the average mass, of a single eukaryotic cell is times greater than its prokaryotic counterpart (refer to de Duve [57] and Milo and Phillips [58]). Hence, the total number of prokaryotic cells on Earth may be approximately three orders of magnitude higher than the equivalent for eukaryotic cells; the relevance of this inference will shortly become apparent.
The total number of prokaryotes is , and the mean number of prokaryotes with which a single prokaryote can communicate is ; we shall conservatively assume that prokaryotes convey information to merely their immediate neighbors. Therefore, the total number of communicating pairs that could exist in principle is . However, of the total population of prokaryotes, only a fraction (which may perhaps approach unity in some settings) are active and taking part in molecular communication at an arbitrary moment in time. The global rate of information transmission attributable to the prokaryotic realm (denoted by ) is accordingly estimated to be
where represents the information transmission rate between a characteristic pair of prokaryotes, conventionally expressed in terms of the mutual information [59]. It is reasonable to concentrate on biofilms henceforth, since a near-majority of all prokaryotes (circa 40–) on our planet are presumed to be residents of biofilms [60].
We specify [60,61] (this value is constructed by taking the estimate of cells from Flemming and Wuertz [60], and multiplying it with a biofilm fraction of ), along with ([62], Figure 1D) (this choice is identical to the number of nearest neighbors manifested in the efficient method of hexagonal close packing in two dimensions), for prokaryotes occurring in biofilms, although the latter could be enhanced under optimal conditions [63,64]. Reliable constraints on do not seem to be available, especially for the substantial deep biosphere, which remains scarcely investigated. We will suppose that of all cells are active at any moment [65,66] (in actuality, even dormant microbes (in the form of spores) are capable of information sensing, and responding to environmental signals [67], but we neglect their relative contribution here), and that the above subset of active cells participate in signaling and communication merely around of the time; the latter may constitute a strongly conservative assumption in some settings (consult, e.g., Larkin et al. [62]). The preceding fiducial values collectively translate to selecting .
Finally, we turn our attention to for prokaryotes. Mattingly et al. [68] tackled Escherichia coli chemotaxis via experiments and theoretical modeling, and determined that information could be acquired at rates up to bits/s, but specifically in environments with shallow chemical gradients (length scales of mm to cm). A theoretical study of E. coli chemotaxis by Tostevin and Ten Wolde [69] arrived at a similar rate, but entailed simplifying assumptions like Gaussian white noise. In a clearly different context, wherein the goal was to effectuate an experimental molecular communication system with an optical-to-chemical signal converter using E. coli, Grebenstein et al. [70] reported an information transmission rate close to bits/s.
On the other hand, E. coli engineered to transmit data (i.e., functioning as molecular communication systems) via modulation schemes such as “time-elapse communication” have been shown to yield information transmission rates of bits/s [71]. Although we focus strictly on intercellular signaling, we remark in passing that intracellular signaling via the ethylene signal transduction pathway in the well-known plant Arabidopsis thaliana might also exhibit information transmission rates of this order. On the basis of this discussion, we adopt a communication rate of bits/s; however, we caution that could be even smaller for certain prokaryotes.
On substituting the prior values into (1), we obtain a global information transmission rate of bits/s for the modern biosphere. This rate, at first glimpse, is many orders of magnitude smaller than bits/s, which is an optimistic estimate for the proposed computational speed of the biosphere [72]. An approximate analog for the technosphere may be global computing power, which is anticipated to potentially reach FLOPS (floating point operations per second) in 2030 if artificial intelligence (AI) computing is taken into account, as per the Intelligent World 2030 report from Huawei, e.g., [73] (https://www.huawei.com/uk/giv, accessed on 18 July 2023), and other informal studies (https://www.diskmfr.com/this-article-is-worth-reading-about-the-computing-power/, accessed on 18 July 2023).
The discrepancy between and the above computational speed of the biosphere is explained by the fact that Landenmark et al. [72] were primarily focused toward quantifying the transcription rate of nucleotides as a proxy for computation, whereas our emphasis is manifestly on information shared between agents via communication channels. An emphasis on information transfer foregrounds the networked nature of the biosphere’s dynamics, which is arguably as important as analyzing the typically self-contained nature of individual DNA replication events.
At this juncture, some key caveats pertaining to (1) are necessary, which also apply to the subsequent (2), albeit to a lesser degree. First, we reiterate that (1) comprises an order-of-magnitude approach that does not reflect, among other effects, spatial heterogeneity and temporal evolution. Second, we see that (1) has a linear dependence on four factors, and each has some uncertainty associated with them. The degree of variation attributable to and is not substantial because biofilms have been widely scrutinized, but and are not tightly constrained. However, as the scaling is linear (and not a higher power-law exponent), our results may not be greatly affected if the estimates provided for these variables are not incorrect by orders of magnitude.
Before moving onward, a brief comment on eukaryotes is warranted. We would need to replace all the parameters in (1) with their eukaryotic equivalents (labeled by the subscript ‘e’), in the case of intercellular communication in eukaryotes. However, because is expected, as outlined earlier in Section 2, there are far more prokaryotes than eukaryotes. Hence, if all other factors are held equal in (1), the information transmission between prokaryotes would dominate that of eukaryotes. In other words, may only be attainable when the remaining variables in (1) are collectively several orders of magnitude higher for eukaryotic cells relative to prokaryotes; however, exact or even approximate values for eukaryotes are not easy to discern. Even if this scenario (of ) were to be applicable, it is still justified to interpret as a plausible lower bound for the information transmission rate associated with the biosphere, because the contribution from eukaryotes is not included by definition in .
3. Information Transmission in the Technosphere
In terms of the evolution of informational dynamics of the biosphere, eukaryotes have played a progressively pivotal role. In particular, one eukaryotic species has shaped Earth profoundly, to wit, Homo sapiens. This pivot to highlighting humans is reminiscent of the comparison of biomass and human-made mass; the latter potentially exceeded the former in the year 2020 [45]. The technosphere engendered by anthropogenic activity has witnessed tremendous growth [42], especially so in the Anthropocene [43,44]. If we consider humans as the “units” in lieu of prokaryotes and introduce the subscript ‘h’, the analogue of (1) for human-to-human communication is duly expressible as
We input for the current human population. Next, the information transmission rate for 17 human languages drawn from 9 language families was estimated by Coupé et al. [74] to equal bits/s. The majority of human-to-human communication likely involves small groups, although a fraction of interactions (e.g., public events) are much larger; we adopt an average value of drawing on these considerations. Finally, as most humans are anticipated to actively communicate for a few hours per day, we will select . After plugging these choices in (2), we obtain a global information transmission rate of bits/s. On comparison with , we notice that is over 12 orders of magnitude lower.
However, an inherent subtlety vis-à-vis the above discussion merits explication. The technosphere in the Anthropocene evinces escalating information flow [42], which extends far beyond direct human-to-human communication (tackled in the prior paragraph) and encompasses the Internet, among other communication channels [75]. The advent of the Internet of Things (IoT), entailing Machine-To-Machine (M2M) connections, is swiftly accelerating this trend [76,77,78], with M2M connections probably comprising half of the total in 2022–2023 [79] (https://www.cisco.com/c/en/us/solutions/collateral/executive-perspectives/annual-internet-report/white-paper-c11-741490.html, accessed on 18 July 2023). Hence, it is worth assessing the global rate of information transmission via human-made networks (denoted by ); in this context, Internet traffic can be singled out because of its pace of development.
The Internet traffic consists of various components, some of which (e.g., mobile networks) are expanding at faster rates than others. We employ consumer Internet Protocol (IP) traffic as the proxy. In Cisco [79], the worldwide IP traffic was estimated to be bytes per year, which translates to . The projected growth rate in Cisco [79] was per year during the time frame of 2017 to 2022, which is equivalent to an e-folding timescale of years. Interestingly, the above growth rate displays excellent agreement with the telecommunication growth rate of in the period of 1986 to 2007 determined in a comprehensive analysis by Hilbert and López [75], after tracking 60 analog and digital technologies across this time span.
While the above paragraph indicates that a growth rate of approximately might be a reasonable assumption, we emphasize that this rate can experience either an increase or decrease over the span of decades. For example, technological breakthroughs could facilitate a boost, certain natural or artificial disasters that destroy infrastructure may lower the growth rate, while other disasters like the COVID-19 pandemic (e.g., [80,81]) can elevate it. Therefore, the ensuing extrapolations on multi-decadal timescales must be interpreted with appropriate caution.
By combining the above data concerning the 2017 IP traffic with the past and expected growth rate, we have
where represents the calendar year. Note, however, that the model is clearly predicated upon the key assumption of sustained exponential growth, which might not be realistic as a consequence of technological, socioeconomic, or ecological factors, along the lines articulated in the prior paragraph.
It is worth recognizing that is also subject to evolution, albeit on geological timescales, owing to which we treat it as effectively constant. However, if we were to travel a few Gyr into the past [82] or Gyr into the future [83], the net primary productivity of Earth would be conceivably a few percent of the current value, intimating that might be proportionally diminished. A precise estimate is difficult because the total biomass is not adequately constrained in those eras; current studies suggest that it may have been at least an order of magnitude lower a few Gyr ago relative to the present day [84].
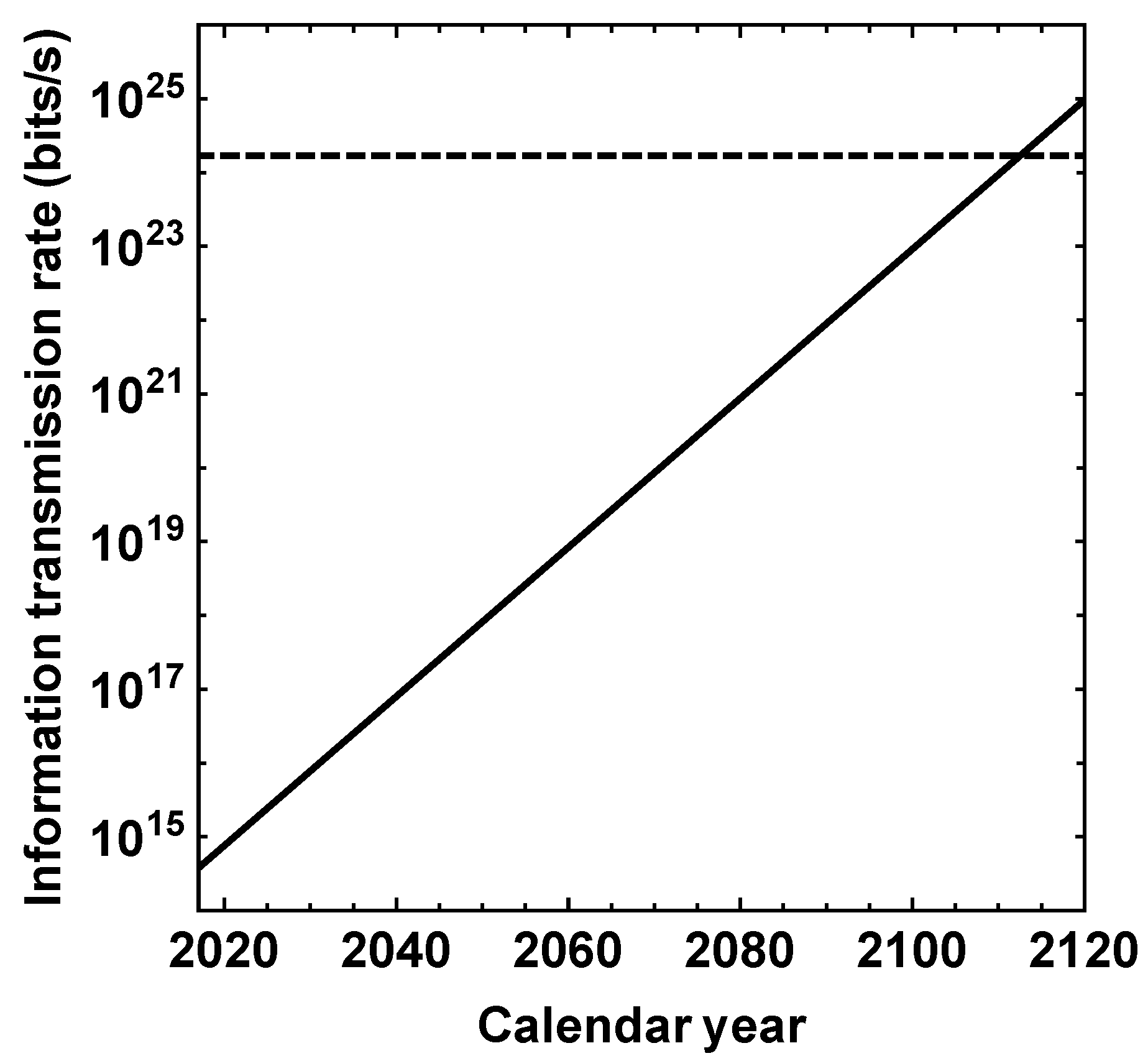
As per the above ansatz, the value of in 2022 is orders of magnitude smaller than our previous estimate for . If (3) still remains valid in the future, then is forecast to exceed in the vicinity of the year 2113 (i.e., about 90 years from now), as evidenced by Figure 1. It is intriguing that the information content (measured in bits) of the technosphere may surpass that stored in DNA (viz., the biosphere) approximately 100 years in the future [46], which is nearly equal to the timescale calculated herein.
4. Discussion and Conclusions
Our analysis, while indubitably simple, yields many crucial implications. First and foremost, the global rate of information transmission in the biosphere might be potentially orders of magnitude higher compared to the technosphere in 2022 (in contrast, the power requirements for communication in the biosphere need not be commensurately higher than the equivalent for the technosphere, given that the thermodynamic efficiency of cellular computations is orders of magnitude greater relative to electronic computing devices [85]). In turn, this prediction indicates that the biosphere is significantly more “communication-rich”, broadly speaking, with respect to the technosphere, although this status quo may undergo transformation in less than a century if the latter exhibits continuous exponential growth due to rapid technological advancements.
Second, this work foregrounds the informational aspects of life, and demonstrates that Earth’s complex biosphere is distinguished by substantial information flow. Hence, traditional yardsticks such as energy (e.g., [86,87]) or energy rate density [88,89] utilized to categorize complex systems may be supplemented by information-centric metrics like the information transmission rate employed herein. For instance, the Kardashev scale in astrobiology [90], which classifies putative technological species (including humans) based on their energy consumption (i.e., power), could be complemented by a scheme centered on the information transmission rate, or alternative measures of information content [91].
Lastly, this line of inquiry opens up novel vistas for further research. To conduct an apples-to-apples comparison, we have restricted ourselves to assessing the global information transmission rates entailing communication. It is feasible to dig deeper into the properties of the underlying information networks and systematically analyze their topology, for example, to ascertain whether they are scale-free, which is ostensibly the case [92]. By the same token, in lieu of canonical (mutual) information, state-of-the-art quantitative concepts endowed with additional nuance such as semantic information [93] or functional information [94] could be harnessed.
Interestingly, while our planet’s biosphere has maintained a mostly stable state—albeit punctuated by evolutionary radiations and mass extinctions [95,96,97,98,99]—over the span of Gyr [100,101,102,103,104,105], the physical facets of the technosphere (engendered by humans) witnessed explosive growth in the Anthropocene [42], to wit, on a timescale of years. In a similar vein, our work suggests that (embodying the informational aspects of the technosphere) is roughly experiencing exponential growth currently, which might potentially lead to this quantity approaching and/or overtaking is less than a century from now. Understanding the information architecture of Earth’s biosphere and technosphere (e.g., relative amounts of semantic and syntactic information) may enable us to go beyond the coarse-grained metric of the global information transmission rate introduced in this paper, and thereby illuminate the differences between the biosphere and technosphere (and their trajectories). This point might prove to be important, as the detrimental effects of the Anthropocene, vis-à-vis long term negative impacts, may conceivably fall more strongly on the technosphere compared to the biosphere.
We round off our exposition by reiterating the major caveats: some of the model parameters are poorly constrained, and this simple order-of-magnitude treatment may not account for certain vital processes. Hence, our approach should be viewed as heuristic, and as a preliminary foray into this subject that can, in principle, facilitate and/or stimulate subsequent research. Of the multiple parameters in our approach, we highlight the need to perform in-depth analyses of the fraction of time that cells communicate in biofilms and the average information transmission rate between each pair of cells in such environments (as mentioned in Section 2). Constraining these variables will help us gain a more reliable understanding of the magnitude of .
Nonetheless, in spite of these limitations, it seems credible that our central conclusion is robust, namely, that information transmission via communication in the biosphere is many orders of magnitude higher than the current technosphere. If this proposition is indeed correct, the biosphere apparently comprises the dominant component of communication when we seek to either comprehend the present or gaze backward in time, but the future trajectory of this phenomenon might be indelibly sculpted by the technosphere in the Anthropocene and beyond.
Author Contributions
Conceptualization, M.L., A.F. and A.B.; methodology, M.L.; formal analysis, M.L.; writing—original draft preparation, M.L.; writing—review and editing, A.F. and A.B.; supervision, A.F. and A.B.; project administration, M.L.; funding acquisition, A.F. All authors have read and agreed to the published version of the manuscript.
Funding
The research undertaken by M.L. and A.F. was funded by the NASA Exobiology program under the grant 80NSSC22K1009. M.L. acknowledges fiscal support from the Florida Tech Open Access Subvention Fund.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
M.L. is grateful to Sandeep Choubey and Chris McKay for perspicacious comments regarding a cognate manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schrödinger, E. What Is Life? The Physical Aspect of the Living Cell; Cambridge University Press: Cambridge, UK, 1944. [Google Scholar]
- Conrad, M. Cross-scale information processing in evolution, development and intelligence. BioSystems 1996, 38, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.M.; Szathmary, E. The Origins of Life: From the Birth of Life to the Origin of Language; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Gatenby, R.A.; Frieden, B.R. Information Theory in Living Systems, Methods, Applications, and Challenges. Bull. Math. Biol. 2007, 69, 635–657. [Google Scholar] [CrossRef]
- Nurse, P. Life, logic and information. Nature 2008, 454, 424–426. [Google Scholar] [CrossRef]
- Krakauer, D.C. Darwinian demons, evolutionary complexity, and information maximization. Chaos 2011, 21, 037110. [Google Scholar] [CrossRef]
- Adami, C. The use of information theory in evolutionary biology. Ann. N. Y. Acad. Sci. 2012, 1256, 49–65. [Google Scholar] [CrossRef]
- Zenil, H. (Ed.) A Computable Universe: Understanding and Exploring Nature as Computation; World Scientific: Singapore, 2013. [Google Scholar]
- Davies, P.C.W.; Rieper, E.; Tuszynski, J.A. Self-organization and entropy reduction in a living cell. Biosystems 2013, 111, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Farnsworth, K.D.; Nelson, J.; Gershenson, C. Living is Information Processing: From Molecules to Global Systems. Acta Biotheor. 2013, 61, 203–222. [Google Scholar] [CrossRef]
- Davies, P.C.W.; Imari Walker, S. The hidden simplicity of biology. Rep. Prog. Phys. 2016, 79, 102601. [Google Scholar] [CrossRef] [PubMed]
- Lan, G.; Tu, Y. Information processing in bacteria: Memory, computation, and statistical physics: A key issues review. Rep. Prog. Phys. 2016, 79, 052601. [Google Scholar] [CrossRef]
- Tkačik, G.; Bialek, W. Information Processing in Living Systems. Annu. Rev. Condens. Matter Phys. 2016, 7, 89–117. [Google Scholar] [CrossRef]
- Uda, S. Application of information theory in systems biology. Biophys. Rev. 2020, 12, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Scharf, C. The Ascent of Information: Books, Bits, Genes, Machines, and Life’s Unending Algorithm; Riverhead Books: New York, NY, USA, 2021. [Google Scholar]
- Fields, C.; Levin, M. Metabolic limits on classical information processing by biological cells. BioSystems 2021, 209, 104513. [Google Scholar] [CrossRef]
- Lingam, M. Theoretical Constraints Imposed by Gradient Detection and Dispersal on Microbial Size in Astrobiological Environments. Astrobiology 2021, 21, 813–830. [Google Scholar] [CrossRef]
- Lyon, P.; Keijzer, F.; Arendt, D.; Levin, M. Reframing cognition: Getting down to biological basics. Philos. Trans. R. Soc. B 2021, 376, 20190750. [Google Scholar] [CrossRef]
- Timsit, Y.; Grégoire, S.P. Towards the Idea of Molecular Brains. Int. J. Mol. Sci. 2021, 22, 11868. [Google Scholar] [CrossRef]
- Frank, A.; Grinspoon, D.; Walker, S. Intelligence as a planetary scale process. Int. J. Astrobiol. 2022, 21, 47–61. [Google Scholar] [CrossRef]
- Raymond, R.C. Communications, Entropy, and Life. Am. Sci. 1950, 38, 273–278. [Google Scholar]
- Loewenstein, W.R. The Touchstone of Life: Molecular Information, Cell Communication, and the Foundations of Life; Oxford University Press: Oxford, UK, 1999. [Google Scholar]
- Fels, D. Cellular Communication through Light. PLoS ONE 2009, 4, e5086. [Google Scholar] [CrossRef]
- Donaldson-Matasci, M.C.; Bergstrom, C.T.; Lachmann, M. The fitness value of information. Oikos 2010, 119, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Benenson, Y. Biomolecular computing systems: Principles, progress and potential. Nat. Rev. Genet. 2012, 13, 455–468. [Google Scholar] [CrossRef]
- Baluška, F.; Levin, M. On Having No Head: Cognition throughout Biological Systems. Front. Psychol. 2016, 7, 902. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.B., Jr. Biological information systems: Evolution as cognition-based information management. Prog. Biophys. Mol. Biol. 2018, 134, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.B., Jr.; Torday, J.S.; Baluška, F. Biological evolution as defense of ‘self’. Prog. Biophys. Mol. Biol. 2019, 142, 54–74. [Google Scholar] [CrossRef] [PubMed]
- Magarini, M.; Stano, P. Synthetic Cells Engaged in Molecular Communication: An Opportunity for Modelling Shannon- and Semantic-Information in the Chemical Domain. Front. Commun. Netw. 2021, 2, 724597. [Google Scholar] [CrossRef]
- Shapiro, J.A. All living cells are cognitive. Biochem. Biophys. Res. Commun. 2021, 564, 134–149. [Google Scholar] [CrossRef]
- Egan, M.; Kuscu, M.; Barros, M.T.; Booth, M.; Llopis-Lorente, A.; Magarini, M.; Martins, D.P.; Schäfer, M.; Stano, P. Toward Interdisciplinary Synergies in Molecular Communications: Perspectives from Synthetic Biology, Nanotechnology, Communications Engineering and Philosophy of Science. Life 2023, 13, 208. [Google Scholar] [CrossRef]
- Margulis, L.; Sagan, D. Microcosmos: Four Billion Years of Microbial Evolution; University of California Press: Berkeley, CA, USA, 1997. [Google Scholar]
- Villarreal, L.P.; Witzany, G. That is life: Communicating RNA networks from viruses and cells in continuous interaction. Ann. N. Y. Acad. Sci. 2019, 1447, 5–20. [Google Scholar] [CrossRef]
- Witzany, G. What is Life? Front. Astron. Space Sci. 2020, 7, 7. [Google Scholar] [CrossRef]
- Shapiro, J.A. Thinking about bacterial populations as multicellular organisms. Annu. Rev. Microbiol. 1998, 52, 81–104. [Google Scholar] [CrossRef]
- Ben-Jacob, E.; Cohen, I.; Levine, H. Cooperative self-organization of microorganisms. Adv. Phys. 2000, 49, 395–554. [Google Scholar] [CrossRef]
- Crespi, B.J. The evolution of social behavior in microorganisms. Trends Ecol. Evol. 2001, 16, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Ben-Jacob, E. Learning from Bacteria about Natural Information Processing. Ann. N. Y. Acad. Sci. 2009, 1178, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Lyon, P. The cognitive cell: Bacterial behavior reconsidered. Front. Microbiol. 2015, 6, 264. [Google Scholar] [CrossRef] [PubMed]
- Levin, M. The Computational Boundary of a “Self”: Developmental Bioelectricity Drives Multicellularity and Scale-Free Cognition. Front. Psychol. 2019, 10, 2688. [Google Scholar] [CrossRef]
- Vernadsky, V.I. The Biosphere; Copernicus Books: New York, NY, USA, 1998. [Google Scholar] [CrossRef]
- Zalasiewicz, J.; Williams, M.; Waters, C.N.; Barnosky, A.D.; Palmesino, J.; Rönnskog, A.S.; Edgeworth, M.; Neal, C.; Cearreta, A.; Ellis, E.C.; et al. Scale and diversity of the physical technosphere: A geological perspective. Anthr. Rev. 2017, 4, 9–22. [Google Scholar] [CrossRef]
- Lewis, S.L.; Maslin, M.A. The Human Planet: How We Created the Anthropocene; Yale University Press: New Haven, CT, USA, 2018. [Google Scholar]
- Thomas, J.A.; Williams, M.; Zalasiewicz, J. The Anthropocene: A Multidisciplinary Approach; John Wiley & Sons: Hoboken, NJ, USA, 2020. [Google Scholar]
- Elhacham, E.; Ben-Uri, L.; Grozovski, J.; Bar-On, Y.M.; Milo, R. Global human-made mass exceeds all living biomass. Nature 2020, 588, 442–444. [Google Scholar] [CrossRef]
- Gillings, M.R.; Hilbert, M.; Kemp, D.J. Information in the Biosphere: Biological and Digital Worlds. Trends Ecol. Evol. 2016, 31, 180–189. [Google Scholar] [CrossRef]
- Schulze-Makuch, D.; Irwin, L.N. Life in the Universe: Expectations and Constraints, 3rd ed.; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Lingam, M.; Loeb, A. Colloquium: Physical constraints for the evolution of life on exoplanets. Rev. Mod. Phys. 2019, 91, 021002. [Google Scholar] [CrossRef]
- Cockell, C.S. Astrobiology: Understanding Life in the Universe, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2020. [Google Scholar]
- Lingam, M.; Loeb, A. Life in the Cosmos: From Biosignatures to Technosignatures; Harvard University Press: Cambridge, UK, 2021. [Google Scholar]
- Drake, F.D. The Radio Search for Intelligent Extraterrestrial Life. In Current Aspects of Exobiology; Mamikunian, G., Briggs, M.H., Eds.; Pergamon Press: Oxford, UK, 1965; pp. 323–345. [Google Scholar]
- Lingam, M.; Loeb, A. Relative Likelihood of Success in the Search for Primitive versus Intelligent Extraterrestrial Life. Astrobiology 2019, 19, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.T.; Haqq-Misra, J.; Frank, A.; Kopparapu, R.; Lingam, M.; Sheikh, S.Z. The Case for Technosignatures: Why They May Be Abundant, Long-lived, Highly Detectable, and Unambiguous. Astrophys. J. Lett. 2022, 927, L30. [Google Scholar] [CrossRef]
- Nakano, T.; Eckford, A.W.; Haraguchi, T. Molecular Communication; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Phillips, R.; Kondev, J.; Theriot, J.; Garcia, H.G. Physical Biology of the Cell, 2nd ed.; Garland Science: New York, NY, USA, 2013. [Google Scholar]
- Bar-On, Y.M.; Phillips, R.; Milo, R. The biomass distribution on Earth. Proc. Natl. Acad. Sci. USA 2018, 115, 6506–6511. [Google Scholar] [CrossRef]
- de Duve, C. The Birth of Complex Cells. Sci. Am. 1996, 274, 50–57. [Google Scholar] [CrossRef]
- Milo, R.; Phillips, R. Cell Biology by the Numbers; Garland Science: New York, NY, USA, 2016. [Google Scholar]
- Levchenko, A.; Nemenman, I. Cellular noise and information transmission. Curr. Opin. Biotechnol. 2014, 28, 156–164. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wuertz, S. Bacteria and archaea on Earth and their abundance in biofilms. Nat. Rev. Microbiol. 2019, 17, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Whitman, W.B.; Coleman, D.C.; Wiebe, W.J. Prokaryotes: The Unseen Majority. Proc. Natl. Acad. Sci. USA 1998, 95, 6578–6583. [Google Scholar] [CrossRef] [PubMed]
- Larkin, J.W.; Zhai, X.; Kikuchi, K.; Redford, S.E.; Prindle, A.; Liu, J.; Greenfield, S.; Walczak, A.M.; Garcia-Ojalvo, J.; Mugler, A.; et al. Signal Percolation within a Bacterial Community. Cell Syst. 2018, 7, 137–145. [Google Scholar] [CrossRef]
- Lee, D.y.D.; Prindle, A.; Liu, J.; Süel, G.M. SnapShot: Electrochemical Communication in Biofilms. Cell 2017, 170, 214.e1. [Google Scholar] [CrossRef] [PubMed]
- Manna, S.; Ghanty, C.; Baindara, P.; Barik, T.K.R.; Mandal, S.M. Electrochemical communication in biofilm of bacterial community. J. Basic Microbiol. 2020, 60, 819–827. [Google Scholar]
- Jones, S.E.; Lennon, J.T. Dormancy contributes to the maintenance of microbial diversity. Proc. Natl. Acad. Sci. USA 2010, 107, 5881–5886. [Google Scholar] [CrossRef]
- Rittershaus, E.S.C.; Baek, S.H.; Sassetti, C.M. The Normalcy of Dormancy: Common Themes in Microbial Quiescence. Cell Host Microbe 2013, 13, 643–651. [Google Scholar] [CrossRef]
- Kikuchi, K.; Galera-Laporta, L.; Weatherwax, C.; Lam, J.Y.; Moon, E.C.; Theodorakis, E.A.; Garcia-Ojalvo, J.; Süel, G.M. Electrochemical potential enables dormant spores to integrate environmental signals. Science 2022, 378, 43–49. [Google Scholar] [CrossRef]
- Mattingly, H.H.; Kamino, K.; Machta, B.B.; Emonet, T. Escherichia coli chemotaxis is information limited. Nat. Phys. 2021, 17, 1426–1431. [Google Scholar] [CrossRef] [PubMed]
- Tostevin, F.; Ten Wolde, P.R. Mutual Information between Input and Output Trajectories of Biochemical Networks. Phys. Rev. Lett. 2009, 102, 218101. [Google Scholar] [CrossRef]
- Grebenstein, L.; Kirchner, J.; Peixoto, R.S.; Zimmermann, W.; Irnstorfer, F.; Wicke, W.; Ahmadzadeh, A.; Jamali, V.; Fischer, G.; Weigel, R.; et al. Biological Optical-to-Chemical Signal Conversion Interface: A Small-Scale Modulator for Molecular Communications. IEEE Trans. Nanobiosci. 2018, 18, 31–42. [Google Scholar] [CrossRef]
- Krishnaswamy, B.; Austin, C.M.; Bardill, J.P.; Russakow, D.; Holst, G.L.; Hammer, B.K.; Forest, C.R.; Sivakumar, R. Time-Elapse Communication: Bacterial Communication on a Microfluidic Chip. IEEE Trans. Commun. 2013, 61, 5139–5151. [Google Scholar] [CrossRef]
- Landenmark, H.K.E.; Forgan, D.H.; Cockell, C.S. An Estimate of the Total DNA in the Biosphere. PLoS Biol. 2015, 13, e1002168. [Google Scholar] [CrossRef] [PubMed]
- Mei, J.; Wei, M.; Sun, Y.; Li, J.; Zhou, G.; Zhang, X. The Architecture of Computing Power Network Towards Federated Learning: Paradigms and Perspectives. In Proceedings of the 2023 IEEE International Symposium on Broadband Multimedia Systems and Broadcasting (BMSB), Beijing, China, 14–16 June 2023; pp. 1–6. [Google Scholar] [CrossRef]
- Coupé, C.; Oh, Y.; Dediu, D.; Pellegrino, F. Different languages, similar encoding efficiency: Comparable information rates across the human communicative niche. Sci. Adv. 2019, 5, eaaw2594. [Google Scholar] [CrossRef] [PubMed]
- Hilbert, M.; López, P. The World’s Technological Capacity to Store, Communicate, and Compute Information. Science 2011, 332, 60–65. [Google Scholar] [CrossRef]
- Atzori, L.; Iera, A.; Morabito, G. The Internet of Things: A survey. Comput. Netw. 2010, 54, 2787–2805. [Google Scholar] [CrossRef]
- Holler, J.; Tsiatsis, V.; Mulligan, C.; Karnouskos, S.; Avesand, S.; Boyle, D. Internet of Things; Academic Press: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Greengard, S. The Internet of Things; The MIT Press: Cambridge, UK, 2021. [Google Scholar]
- Cisco. Cisco Visual Networking Index: Forecast and Trends, 2017–2022. 2019. Available online: https://twiki.cern.ch/twiki/pub/HEPIX/TechwatchNetwork/HtwNetworkDocuments/white-paper-c11-741490.pdf (accessed on 26 August 2023).
- Feldmann, A.; Gasser, O.; Lichtblau, F.; Pujol, E.; Poese, I.; Dietzel, C.; Wagner, D.; Wichtlhuber, M.; Tapiador, J.; Vallina-Rodriguez, N.; et al. A year in lockdown: How the waves of COVID-19 impact internet traffic. Commun. ACM 2021, 64, 101–108. [Google Scholar] [CrossRef]
- Whalley, J.; Stocker, V.; Lehr, W. (Eds.) Beyond the Pandemic? Exploring the Impact of COVID-19 on Telecommunications and the Internet; Emerald Publishing Limited: Bingley, UK, 2023. [Google Scholar] [CrossRef]
- Hao, J.; Knoll, A.H.; Huang, F.; Schieber, J.; Hazen, R.M.; Daniel, I. Cycling phosphorus on the Archean Earth: Part II. Phosphorus limitation on primary production in Archean ecosystems. Geochim. Cosmochim. Acta 2020, 280, 360–377. [Google Scholar] [CrossRef]
- Ozaki, K.; Reinhard, C.T. The future lifespan of Earth’s oxygenated atmosphere. Nat. Geosci. 2021, 14, 138–142. [Google Scholar] [CrossRef]
- McMahon, S.; Parnell, J. The deep history of Earth’s biomass. J. Geol. Soc. 2018, 175, 716–720. [Google Scholar] [CrossRef]
- Kempes, C.P.; Wolpert, D.; Cohen, Z.; Pérez-Mercader, J. The thermodynamic efficiency of computations made in cells across the range of life. Philos. Trans. R. Soc. A 2017, 375, 20160343. [Google Scholar] [CrossRef] [PubMed]
- Smil, V. Energy in Nature and Society: General Energetics of Complex Systems; The MIT Press: Cambridge, UK, 2008. [Google Scholar]
- Smil, V. Energy Transitions: History, Requirements, Prospects; ABC-CLIO, LLC: Santa Barbara, CA, USA, 2010. [Google Scholar]
- Chaisson, E.J. Cosmic Evolution: The Rise of Complexity in Nature; Harvard University Press: Cambridge, UK, 2001. [Google Scholar]
- Chaisson, E.J. Energy rate density as a complexity metric and evolutionary driver. Complexity 2011, 16, 27–40. [Google Scholar] [CrossRef]
- Kardashev, N.S. Transmission of Information by Extraterrestrial Civilizations. Soviet Astron. 1964, 8, 217–221. [Google Scholar]
- Wright, J.T.; Cartier, K.M.S.; Zhao, M.; Jontof-Hutter, D.; Ford, E.B. The Search for Extraterrestrial Civilizations with Large Energy Supplies. IV. The Signatures and Information Content of Transiting Megastructures. Astrophys. J. 2016, 816, 17. [Google Scholar] [CrossRef]
- Kim, H.; Smith, H.B.; Mathis, C.; Raymond, J.; Walker, S.I. Universal scaling across biochemical networks on Earth. Sci. Adv. 2019, 5, eaau0149. [Google Scholar] [CrossRef]
- Kolchinsky, A.; Wolpert, D.H. Semantic information, autonomous agency and non-equilibrium statistical physics. Interface Focus 2018, 8, 20180041. [Google Scholar] [CrossRef]
- Hazen, R.M.; Griffin, P.L.; Carothers, J.M.; Szostak, J.W. Functional information and the emergence of biocomplexity. Proc. Natl. Acad. Sci. USA 2007, 104, 8574–8581. [Google Scholar] [CrossRef]
- Hazen, R.M. The Story of Earth: The First 4.5 Billion Years, from Stardust to Living Planet; Penguin Books: New York, NY, USA, 2012. [Google Scholar]
- Dawkins, R.; Wong, Y. The Ancestor’s Tale: A Pilgrimage to the Dawn of Evolution, 2nd ed.; Mariner Books: Boston, MA, USA, 2016. [Google Scholar]
- Knoll, A.H.; Nowak, M.A. The timetable of evolution. Sci. Adv. 2017, 3, e1603076. [Google Scholar] [CrossRef] [PubMed]
- Benton, M.J. (Ed.) Cowen’s History of Life, 6th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2020. [Google Scholar]
- Knoll, A.H. A Brief History of Earth: Four Billion Years in Eight Chapters; Custom House: New York, NY, USA, 2021. [Google Scholar]
- Kirchner, J.W. The Gaia hypothesis: Can it be tested? Rev. Geophys. 1989, 27, 223–235. [Google Scholar] [CrossRef]
- Caldeira, K.; Kasting, J.F. The life span of the biosphere revisited. Nature 1992, 360, 721–723. [Google Scholar] [CrossRef] [PubMed]
- Lenton, T.M. Gaia and natural selection. Nature 1998, 394, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Free, A.; Barton, N.H. Do evolution and ecology need the Gaia hypothesis? Trends Ecol. Evol. 2007, 22, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Lenton, T.M.; Daines, S.J.; Dyke, J.G.; Nicholson, A.E.; Wilkinson, D.M.; Williams, H.T.P. Selection for Gaia across Multiple Scales. Trends Ecol. Evol. 2018, 33, 633. [Google Scholar] [CrossRef]
- Kasting, J.F. The Goldilocks Planet? How Silicate Weathering Maintains Earth “Just Right”. Elements 2019, 15, 235–240. [Google Scholar] [CrossRef]
Figure 1.
Information transmission rate associated with communication (in bits/s) as a function of the calendar year. The dashed line is the rate estimated for the biosphere (assuming it is roughly constant on short timescales) and the solid line signifies the rate for the technosphere given by (3).
Figure 1.
Information transmission rate associated with communication (in bits/s) as a function of the calendar year. The dashed line is the rate estimated for the biosphere (assuming it is roughly constant on short timescales) and the solid line signifies the rate for the technosphere given by (3).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lingam, M.; Frank, A.; Balbi, A. Planetary Scale Information Transmission in the Biosphere and Technosphere: Limits and Evolution. Life 2023, 13, 1850. https://doi.org/10.3390/life13091850
AMA Style
Lingam M, Frank A, Balbi A. Planetary Scale Information Transmission in the Biosphere and Technosphere: Limits and Evolution. Life. 2023; 13(9):1850. https://doi.org/10.3390/life13091850
Chicago/Turabian StyleLingam, Manasvi, Adam Frank, and Amedeo Balbi. 2023. "Planetary Scale Information Transmission in the Biosphere and Technosphere: Limits and Evolution" Life 13, no. 9: 1850. https://doi.org/10.3390/life13091850
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.